 Jie Wu1
Jie Wu1 Baojun He2Miao Miao3Xibin Han4Hongyan Dai5Heng Dou6
Baojun He2Miao Miao3Xibin Han4Hongyan Dai5Heng Dou6 Yanqiu Li6Xiaoqing Zhang7*
Yanqiu Li6Xiaoqing Zhang7* Guangchuan Wang3*
Guangchuan Wang3*- 1Department of Oncology, The First Affiliated Hospital of Jinzhou Medical University, Jinzhou, China
- 2Department of Clinical Laboratory Medicine, The First Affiliated Hospital of Jinzhou Medical University, Jinzhou, China
- 3Department of Immunology, School of Basic Medical Science, Jinzhou Medical University, Jinzhou, China
- 4Laboratory Animal Center, Jinzhou Medical University, Jinzhou, China
- 5Department of Outpatient PICC, The First Affiliated Hospital of Jinzhou Medical University, Jinzhou, China
- 6Greatest Biopharma Limited Company, Benxi, China
- 7Teaching Center for Basic Medical Experiment, China Medical University, Shenyang, China
The biological macromolecule Nocardia rubra cell-wall skeleton (Nr-CWS) has well-established immune-stimulating and anti-tumor activities. However, the role of Nr-CWS on natural killer (NK) cells remains unclear. Here, we explore the function and related mechanisms of Nr-CWS on NK cells. Using a tumor-bearing model, we show that Nr-CWS has slightly effect on solid tumor. In addition, using a tumor metastasis model, we show that Nr-CWS suppresses the lung metastasis induced by B16F10 melanoma cells in mice, which indicates that Nr-CWS may up-regulate the function of NK cells. Further investigation demonstrated that Nr-CWS can increase the expression of TRAIL and FasL on spleen NK cells from Nr-CWS treated B16F10 tumor metastasis mice. The spleen index and serum levels of TNF-α, IFN-γ, and IL-2 in B16F10 tumor metastasis mice treated with Nr-CWS were significantly increased. In vitro, the studies using purified or sorted NK cells revealed that Nr-CWS increases the expression of CD69, TRAIL, and FasL, decreases the expression of CD27, and enhances NK cell cytotoxicity. The intracellular expression of IFN-γ, TNF-α, perforin (prf), granzyme-B (GrzB), and secreted TNF-α, IFN-γ, IL-6 of the cultured NK cells were significantly increased after treatment with Nr-CWS. Overall, the findings indicate that Nr-CWS could suppress the lung metastasis induced by B16F10 melanoma cells, which may be exerted through its effect on NK cells by promoting NK cell terminal differentiation (CD27lowCD11bhigh), and up-regulating the production of cytokines and cytotoxic molecules.
Introduction
Nocardia rubra is a species of gram-positive, rod-shaped bacteria that is not pathogenic to human [1]. Nocardia rubra cell-wall skeleton (Nr-CWS) was first reported in 1976 [2]. The compositional analysis revealed that Nr-CWS is composed of 0.35% D-mannose, 0.63% D-ribose, 2.60% L-rhamnose monohydrate, 1.83% D-ructose, 5.92% D-glucose, 2.54% D-xylose, 32.36% D-lactose, and 53.76% arabinose. Early investigations have reported that Nr-CWS is with high immuno-stimulating and anti-tumor activities, and could directly stimulate macrophages, CD4+, and CD8+ T cells [2-7]. In combination with chemotherapy or through administration locally at the tumor site, Nr-CWS prolongs the survival of patients with advanced cancers and increase the levels of cytokines, such as IL-1, IL-6, TNF-α and IFN, in tumor tissue [8]. In addition, Nr-CWS can improve the therapeutic efficacy of PD-1/PD-L1 immunosuppressive pathway in patients with high-risk human papillomavirus (HR-HPV) infection and cervical precancerous lesion, cervical intraepithelial neoplasia (CIN), and mouse CT26. WT cancer cell [6, 9]. Our previous study showed that Nr-CWS can promote CD4+ T cells activation, drive Th1 immune response, and enhance CD8+ T cells function [4, 5]. Recently, the application of Nr-CWS products has been approved in China (approval Number S20030009), for cervical cancer treatment. However, whether Nr-CWS is able to modulate the function of NK cells in tumor metastasis is still under elucidation.
NK cells are major players of innate immunity. They respond quickly against various types of pathogens [10] and play critical roles in immuno-surveillance against tumors [11]. It has been well established that NK cells participate in many physiological and pathophysiological processes, including immuno-surveillance against tumors, anti-intracellular pathogen infection, allograft rejection, and pregnancy [12, 13]. NK cells can remove targets through different pathways [14], including the release of cytotoxic molecules (like perforin, granzymes), induction of apoptosis through TRAIL and FasL, and directly killing targets through antibody-dependent cellular cytotoxicity (ADCC) [15]. NK cells are functionally heterogeneous, which are divided into different subgroups of maturation with different levels of cytokines and cytotoxic capacity based on the expression of CD27 and CD11b [16]. Accumulated evidence shows that NK cells can acquire function through a broad spectrum of molecular and cellular mechanisms [11, 15–17]. In this study, using in vitro and in vivo models, we found that Nr-CWS could suppress the lung metastasis of B16F10 melanoma model, which may be through its effect on NK cells by promoting NK cell terminal differentiation (CD27lowCD11bhigh) and up-regulating the production of cytokines and cytotoxic molecules.
Materials and Methods
Reagents
Nr-CWS was supplied by Greatest Biopharma Limited Company (purity>98%). Fluorescein conjugated anti-mouse antibodies APC-NK1.1, FITC-NK1.1, APC/Cy7-CD3ε, PE/Cy7-CD3ε, APC-CD4, PE-CD8α, PerCP/Cy5.5-CD11b, PE-CD27, FITC-TNF-α, and PerCP/Cy5.5-IFN-γ from BioLegend, and PerCP/Cy5.5-CD69, FITC-FasL, PE-TRAIL, PE-Perforin, and FITC-Granzyme-B from eBioscience were used in this study. TUNEL assay kit and MTS Cell proliferation, cytotoxicity or chemosensitivity assays kit (cat # 3580) were obtained from Promega. The LEGENDplex™ Multiplex mouse cytokine kit was provided by BioLegend. EasySep™ mouse NK cell isolation kit was from STEMCELL Technologies Inc. Recombinant murine IL-2 was from PeproTech Inc.
Tumor Models and Nocardia rubra Cell-Wall Skeleton Administration
Female C57BL/6 mice (6–8 weeks, 18–20 g) supplied by Beijing Vital River Laboratory Animal Technology, were maintained in specific pathogen-free animal rooms and treated following the guideline for Care and Use of Laboratory Animals of Jinzhou Medical University. Lewis lung carcinoma (LLC) and B16F10 melanoma cell lines were bought from Cell Bank of Chinese Academy of Sciences in Shanghai. The cells were grown in DMEM medium supplemented with 10% (v/v) fetal bovine serum (FBS), 100 U/ml penicillin and 100 μg/ml streptomycin, in a humidified atmosphere of 5% CO2 at 37°C. LLC cells (5×106/ml in PBS) were subcutaneously injected in the medial axillas of 22 C57BL/6 mice (0.1 ml/mouse) and after establishment of tumor model for 24 h (hrs), mice were divided into two groups randomly (11 mice/group, Nr-CWS group and control group). For experimental metastasis, B16F10 melanoma cells (1×106/ml in PBS) were injected into C57BL/6 mice through tail-vein (0.2 ml/mouse) and the injected mice were divided into two groups randomly 24 h later (4 mice/group, Nr-CWS group, control group). Nr-CWS group were given Nr-CWS by intra-peritoneal injection (10 mg/kg in PBS, once-daily) for 7 or 14 successive days, and the control group were treated with equivalent PBS.
Sample Processing
For the mouse tumor-bearing model (11 mice/group), 5 mice from each group were euthanized with an overdose of isoflurane on day 7 after tumor cell inoculation. Then solid tumors were dissected for analyses of tumor weight and the percentage of tumor-infiltrating lymphocytes. After 14 days, the remained mice (6 mice/group) were euthanized for sampling: the solid tumors were dissected, each tumor sample was sectioned for tumor morphology and tumor cell apoptosis assay. For experimental metastasis model (one mice in control group died during experiment was excluded), pulmonary metastases were assessed by comparing the numbers of metastases on day 14 after tumor cell injection. Blood serum and spleen were obtained from the metastasis model mice. Index for spleen was calculated as previously reported [18].
Analysis of Tumor Morphology by Hematoxylin and Eosin Staining and Tumor Cells Apoptosis by TUNEL
Tumor morphology was analyzed as previously described [19]: in brief, tumor tissues from Nr-CWS treated and control groups (day 14) were fixed using paraformaldehyde, Fixed samples were embedded in wax after dehydration and clearing using a series of ethanol solution and xylene, thin slices (4 μm) were cut and mounted on microscope slides, samples were stained using hematoxylin and eosin stain (H&E) after deparaffinized and rehydration. Finally slides were mounted using mounting medium and analyzed [19].
Tumor cell apoptosis was detected using TUNEL Assay Kit, the ratio of positive cells per specimen was analyzed under a light microscope. For each sample, five most obvious positive staining areas were analyzed [20]. TUNEL positive cells were counted and apoptotic index (AI) was calculated in reference of a previous publication [20].
Analysis of Tumor Infiltrating Lymphocytes
Each tumor sample dissected on day 7 after tumor cell inoculation was digested using trypsin (0.05%) and mashed through moisturized cell strainer (70 μm), then the single-cell suspension was collected and counted. 1×106 cells were taken for staining. The cocktail of antibodies (APC-CD4, PE/Cy7-CD3ε, FITC-NK1.1, and PE-CD8α) was added into the cell suspension and then the stained samples were incubated for 20 min at 4°C. For dead cell exclusion, 5 µl of 7-Aminoactinomycin D (7-AAD) was used for each sample before analysis. Then samples were detected using flow cytometry (FCM) (BD FACS Canto II) and 30,000 events were acquired and analyzed using BD Diva software.
Natural Killer Cell Purification and Cell Culture
Spleens from wild type C57BL/6 mice were mashed through a moisturized cell strainer. NK cell was purified with EasySep™ mouse NK cell isolation kit and its purity was evaluated by FCM. For the cytokines analysis in culture supernatant and NK cell cytotoxicity assay, purified NK cells were subjected to fluorescence-activated cell sorting (BD FACSAriaIII) to exclude other cell contamination using CD3ε and NK1.1 staining. Freshly isolated NK cells or sorted NK cells were seeded in 96-well flat-bottom plate (1×105/well) containing RPMI-1640 complete medium supplemented with 10% FBS, 100 U/ml penicillin and 100 μg/ml streptomycin, 50 μM β-mercaptoethanol, 500 U/ml IL-2, and treated with or without 15 μg/ml Nr-CWS (the optimal dose for splenocytes from our previous experiments) in a humidified atmosphere of 5% CO2 at 37°C. After 24 or 48 h incubation respectively, NK cells were collected for staining. The culture supernatant was collected for multi-cytokines analysis by FCM (BD Canto II).
Analysis of Cell Surface Molecules Expression by Flow Cytometry
For NK cells analysis from in vivo experiment, spleen single-cell suspension were made by mashing whole spleen through a moisturized cell strainer and RBC lysing. For cultured NK cells analysis in vitro, NK cell was purified from spleen single-cell suspension with EasySep™ mouse NK cell isolation kit. NK cells from spleen of Nr-CWS treated and control B16F10 metastasis mice (after 14 days treatment), and the cultured wild type (WT) spleen NK cells incubated with or without Nr-CWS (15 μg/ml) at indicated time-points were collected for analysis of surface molecules expression by FCM. And cells from each sample were suspended in 100 μl staining buffer. A cocktail of antibodies (combination of the fluorescent conjugated antibodies CD3ε, NK1.1, TRAIL, FasL, CD69 or CD3ε, NK1.1, CD11b, and CD27) was used and the stained samples were incubated for 20 min (4°C). After washing with PBS, the stained cells were analyzed by FCM.
Analysis of Intracellular Perforin/Granzyme-B and Cytokines
Purified WT NK cells treated with or without Nr-CWS (15 μg/ml) were collected at indicated time-points. Monensin (BioLegend) was present for the last 5 h before collection. Then, samples were stained with APC-NK1.1 and APC/Cy7-CD3ε antibodies for surface markers. For intracellular staining, samples were fixed and permeabilized after surface staining using Fixation Buffer (cat # 420801) and Intracellular Staining Permeabilization Wash Buffer (10×, cat # 421002). Then, samples were further stained with the cocktail of cytotoxic molecules (PE-Perforin, FITC-Granzyme B) or cytokines (FITC-TNF-α, PerCP/Cy5.5-IFN-γ) antibodies. After intracellular staining and extensive washing, samples were subjected to FCM analysis.
Multiplex Analysis of Cytokines by Flow Cytometry
The cytokines in the mouse serum of metastasis model and WT NK cell culture supernatants were measured using the LEGENDplex™ Multiplex mouse cytokine kit from BioLegend by FCM as we have reported [5]. The serum from Nr-CWS treated (for 14 days) metastasis mice and control mice were collected and stored at −80°C. The supernatants of cultured NK cells treated with or without Nr-CWS (15 μg/ml) were collected at indicated time-points. Cytokines (IL-2, IL-4, IL-5, IL-6, IL-10, IFN-γ and TNF-α) were evaluated in all serum and supernatant samples. Results were analyzed using LEGENDplex software as previously reported [4, 5].
MTS Assay for Natural Killer Cell Cytotoxicity In Vitro
The assay for NK cell cytotoxicity was performed as previously described by MTS assay [19]. Briefly, NK cells sorted from wild type C57BL/6 mice were stimulated with or without Nr-CWS (15 μg/ml) for 24 h and collected as effectors after washing. LLC cells were serviced as targets. The effector NK cells were mixed with LLC cells (5×103/well) at effector:target ratios of 10:1 and 5:1, and incubated in 96-well flat-bottom plate (Corning-Costar) in 100 µl volume for 6 h. At the last 1 hour, 20 µl/well of CellTiter 96® AQueous One Solution Reagent (MTS) was added. After 1 h at 37°C in a humidifi ed, 5% CO2 atmosphere, the optical density (OD) was measured using multiskan spectrum microplate spectrophotometer at 490 nm from triple wells. The cytotoxicity was calculated as previous report [19]:
Statistical Analysis
Data were shown using mean ± standard deviation (SD). The Independent-Samples t Test was used to analyze results in SPSS 16.0 (SPSS, Inc., Chicago, IL, United States). The p-value <0.05 was considered as statistically significant, as shown in the figure legends. Data shown in figures are representative of three independent experiments.
Results
The Weight and Histopathology of Solid Tumors
To confirm the anti-tumor activity of Nr-CWS, C57BL/6 wild type mice were implanted with LLC cells and then treated with either Nr-CWS or PBS control. The results showed that the average tumor weight of the Nr-CWS group was decreased after 7 days treatment when compared to the control group (p < 0.05). However, it was comparable after 14 days treatment (p > 0.05) (Figure 1A; Supplementary Figure S1A). The histopathological examination of tumors in the Nr-CWS group (after 14 days treatment) showed larger necrotic areas and more prominent infiltration of inflammatory cells than that in the control group. H&E staining showed the tumor cells were poorly differentiated and heterogeneous, with hyperchromatic nuclei and scant cytoplasm. There were also binucleated or multinucleated cells in the tumor samples. The representative histopathological changes are shown in Supplementary Figures S1B,C.
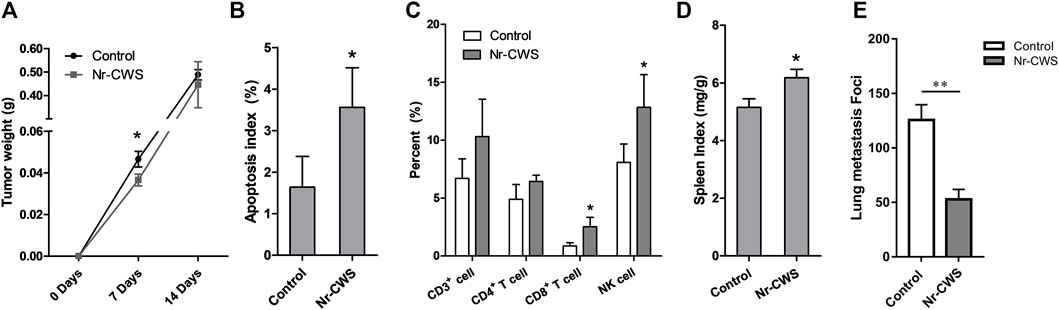
FIGURE 1. Potential enhancement of anti-tumor immune responses by Nr-CWS. (A) The statistical analysis of tumor weights of Nr-CWS treated mice compared with that of control mice at day 7 and 14. (B) The analysis of apoptotic cells in tumor tissues by TUNEL. Histogram showing the apoptotic index of two study groups. (C) Tumor infiltrating lymphocytes were analyzed by FCM. The histogram showed the percentage of CD3+, CD4+ T, CD8+ T, and NK cells in Nr-CWS treated mice and control mice. (D) The spleen index of Nr-CWS treated mice and control mice (after 14 days treatment). (E) Quantification of metastases from mice treated with or without Nr-CWS at day 14 after tail vein injection of B16F10 melanoma cells (0.2 M/mouse). Data are shown as means ± SD; *, p < 0.05; **, p < 0.01.
Analysis of Tumor Cells Apoptosis by TUNEL
The TUNEL System was used for detecting apoptotic cells and apoptotic bodies in situ of solid tumors. The morphological changes of apoptotic cells were observed as follows: apoptotic DNA fragments were stained brown mainly in the condensed nuclei (Supplementary Figure S1D). The apoptotic index was calculated using the percentage of stained brown cells at ×400 magnification. Results showed that Nr-CWS treatment caused more tumor cell apoptosis and higher apoptotic index compared with the control group (p < 0.05, Figure 1B, Supplementary Figure S1D).
Analysis of Tumor Infiltrating Lymphocytes, Spleen Index, and B16F10 Lung Metastasis
For each tumor sample from the tumor-bearing mice after 7 days treatment with or without Nr-CWS, the percentage of infiltrated lymphocytes including CD3+, CD4+ T, CD8+ T, and NK cells, were analyzed by FCM (Figure 1C, Supplementary Figure S1E). The percentage of CD8+ T and NK cells were significantly increased after Nr-CWS treatment (p < 0.05), while the percentage of CD3+ and CD4+ T cells were only slightly up-regulated (p > 0.05) (Figure 1C, Supplementary Figure S1E).
The spleen index of the Nr-CWS treated group was significantly increased after 14 days treatment (p < 0.05), while the spleen index of the Nr-CWS and control group did not show any significant difference (data not shown) after 7 days treatment, as shown in Figure 1D.
To further investigate the role of Nr-CWS in vivo, we adopted the B16F10 melanoma metastasis model. Results showed Nr-CWS treatment can significantly reduce the lung metastasis compared to the control group in Figure 1E, Supplementary Figure S1F (p < 0.01).
Analysis of Natural Killer Cell Purity by Flow Cytometry
The NK cells were purified using EasySep™ mouse NK cell isolation kit based on Magnetic-activated cell sorting (MACS). All living cells (7-AAD negative) were analyzed. The percentage of NK cell in spleen before purification was 2.63 ± 0.31%. After purification, the percentage of NK cell was >80% (81.97 ± 3.40%) as determined by FCM gating on NK1.1+ CD3ε− population (Supplementary Figure S1G). For the cytokines analysis in culture supernatant and NK cell cytotoxicity assay, purified NK cells were subjected to fluorescence-activated cell sorting to exclude other cell contamination using CD3ε and NK1.1 staining. The purity of sorted NK cell was more than 98% (data not shown).
Analysis the Expression of Natural Killer Cell Functional Markers
To assess the effect of Nr-CWS on the functional molecules of NK cells, FCM analysis was performed to determine the expression of functional markers on NK cells gated the population of NK1.1+ CD3ε− cells. The in vivo samples showed that the expression of TRAIL and FasL on splenic NK cell were much higher in the Nr-CWS group than the control group (p < 0.05, Figure 2A, Supplementary Figure S2A). However, CD69 expression and splenic NK cell percentage were comparable between the two groups (p > 0.05, Figures 2A,B). Results of the in vitro experiment (Supplementary Figures S2B,C) showed Nr-CWS can strikingly increase the expression of CD69, TRAIL, and FasL on NK cells after incubation for 24 h (p < 0.01, Figure 2C) and increase CD69 expression after incubation for 48 h (p < 0.01, Figure 2D).
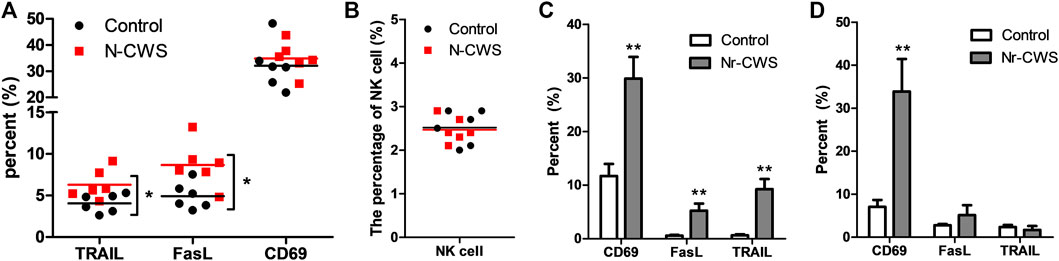
FIGURE 2. Expression of functional molecules of NK cells after Nr-CWS treatment in vivo and in vitro. The graphs showed the percentage of CD69, TRAIL, and FasL positive NK cells in vivo [(A), n = 6] and in vitro [(C), 24 h; (D), 48 h], and the percentage of spleen NK cells in Nr-CWS treated mice and control mice [(B), n = 6]. Data are shown as means ± SD; *, p < 0.05; **, p < 0.01.
Analysis of Perforin/Granzyme-B Expression In Vitro
To investigate the effect of Nr-CWS on NK cell cytotoxicity, the expression of intracellular perforin and granzyme-B of NK cells (gated on NK1.1+ CD3ε−) treated with or without Nr-CWS for 24 or 48 h, were analyzed by FCM. Results showed that Nr-CWS significantly increased the expression of granzyme-B and perforin after incubation for 24 and 48 h, as shown in Figures 3A,B, Supplementary Figures S3A,B.
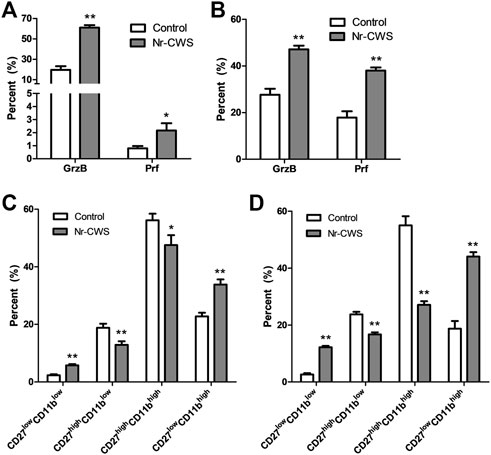
FIGURE 3. Analysis of NK cell subgroups and the expression of perforin and granzyme B in vitro. (A,B) The statistical analyses of the percentage of Prf and GrzB of the Nr-CWS treated and control NK cells by flow cytometry [(A), 24 h; B, 48 h]. (C,D) The statistical analyses of NK cell subgroups based on the expression of CD11b and CD27 by flow cytometry. Graphs showed the statistical analyses of the Nr-CWS treated and control NK cells [(C), 24 h; (D), 48 h]. Data are shown as means ± SD; *, p < 0.05; **, p < 0.01.
Analysis of Natural Killer Cell Subgroups In Vitro
Purified normal spleen NK cells treated with or without Nr-CWS were collected at indicated time-points for subgroups analysis by FCM gating on NK1.1+CD3ε− population. Our data showed that Nr-CWS treatment can significantly up-regulate the percentage of CD27lowCD11bhigh and CD27lowCD11blow cells, and down-regulate the percentage of CD27highCD11blow and CD27highCD11bhigh cells after incubation for 24 and 48 h, as shown in Figures 3C,D and Supplementary Figures S3C,D.
Analysis of Intracellular Cytokines
Purified normal spleen NK cells treated with or without Nr-CWS were collected at indicated time-points for intracellular cytokine analysis by FCM gating on NK1.1+CD3ε− population. Results showed that Nr-CWS can stimulate NK cells to produce significantly more IFN-γ and TNF-α after treatment for 24 and 48 h (p < 0.01, Figures 4A,B, Supplementary Figures S4A,B).
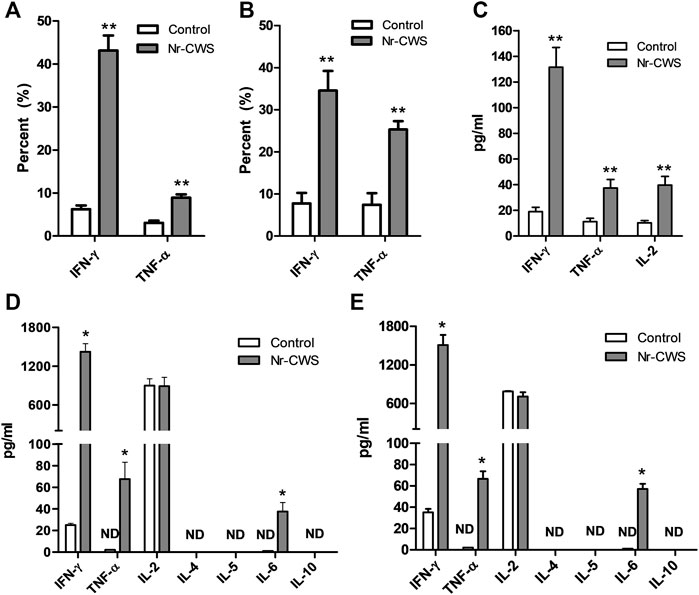
FIGURE 4. Analysis of cytokines. (A,B) Quantification of the expression of IFN-γ and TNF-α of NK cells treated with or without Nr-CWS by flow cytometry. The statistical analyses of the expression of IFN-γ and TNF-α of NK cells treated with or without Nr-CWS in vitro [(A), 24 h; (B), 48 h]. (C) The statistical analyses of the cytokines (IFN-γ, TNF-α, IL-2) in serum after Nr-CWS treatment in vivo. (D,E) The statistical analyses of secreted cytokines (IL-2, IL-4, IL-5, IL-6, IL-10, TNF-α, and IFN-γ) in the supernatants of cultured NK cells treated with or without Nr-CWS in vitro [(D), 24 h; (E), 48 h]. Data are shown as means ± SD; ND, not detected; *, p < 0.05; **, p < 0.01.
Analysis of Secreted Cytokines in Serum and Natural Killer Cell Culture Supernatant
The levels of cytokines in the supernatant and serum from NK cell culture and animals treated with or without Nr-CWS were analyzed by FCM using LEGENDplex™. The serum levels of IL-2, IFN-γ, and TNF-α were significantly higher after Nr-CWS treatment in vivo (p < 0.01, Figure 4C). Results of cytokines in the cultured supernatants showed Nr-CWS significantly increased the production of IFN-γ and TNF-α after treatment for 24 and 48 h in vitro. In contrast, the level of IL-2 was comparable between the two groups (p > 0.05). Nr-CWS can drive the expression of IL-6, which were 37.51 ± 8.32 and 57.16 ± 4.80 pg/ml after incubation for 24 and 48 h in vitro, whereas it was not detectable (ND) in the control group, as shown in Figures 4D,E.
Natural Killer Cells Mediated Cytotoxicity In Vitro
To measure the cytotoxic activity, sorted NK cells were treated with or without Nr-CWS for 24 h. Then the NK cells were washed and the number was accounted, and mixed with LLC cells at indicated ratios for 6 h. The cytotoxic activity was measured by MTS assay. Results showed that Nr-CWS can strikingly increase NK cell-mediated cytotoxicity at both effector:target ratios of 10:1 and 5:1 (p < 0.05, Figure 5).
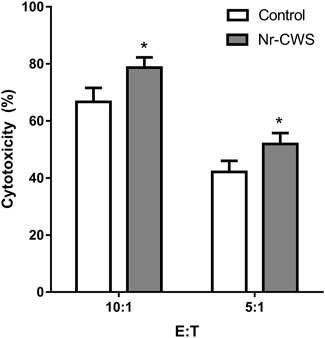
FIGURE 5. Cytotoxicity of NK cells treated with Nr-CWS against LLC cells. Histogram showed the statistical analysis of NK cell cytotoxicity in the Nr-CWS group versus that in the control group at the indicated E:T ratios. Data shown are representative of at least three independent experiments, and calculated data are shown as means ± SD; *, p < 0.05.
Discussion
NK cells, belonging to the innate immune system, unlike T and B cells, play essential roles in the anti-virus or anti-cancer responses by directly killing target cells through rapidly releasing cytotoxic molecules and cytokines, or transduction of apoptosis signaling using surface receptors without prior sensitization [21]. NK cells are efficient and timely to maintain immune homeostasis, especially in early stage of virus-infection and tumor development. In consideration of the compositions and immune-stimulatory function of Nr-CWS we and other groups have reported earlier [3-7], we further explored the immune-regulatory role of Nr-CWS on NK cell against cancer.
In our study, we first explored the anti-tumor effects of Nr-CWS using the LLC tumor-bearing C57BL/6 mice model in vivo. The results showed that there is no apparent effect of Nr-CWS on solid tumor and there is no significant difference of tumor weight at the late stage of tumor growth after Nr-CWS treatment, although Nr-CWS can slightly suppress the tumor growth at the early stage (treated for 7 days; Figure 1A, Supplementary Figure S1A) and induces tumor cell apoptosis (treated for 14 days; Figure 1B, Supplementary Figures S1B,D). It is interesting that the Nr-CWS treated tumor-bearing mice showed significantly up-regulated percentages of tumor infiltrating CD8+ T and NK cells at the early stage (treated for 7 days; Figure 1C, Supplementary Figure S1E), and higher spleen index (treated for 14 days; Figure 1D). A previous study indicated that NK cells can inhibit tumor growth once they enter a tumor site [22] and strikingly delay tumor formation in a model of spontaneous mammary carcinoma at the earliest stages of carcinogenesis rather than late stages in a perforin-dependent manner [23, 24]. Our findings suggest that Nr-CWS may prominently enhance NK cell function. To test this possibility, the B16F10 melanoma metastasis model, which is the most commonly used metastatic melanoma model for preclinical studies was adopted to evaluate the effect of Nr-CWS in vivo [25]. As expected, our result showed that Nr-CWS significantly reduced the B16F10 melanoma lung metastasis (Figure 1E, Supplementary Figure S1F), which may be exerted through its effect on NK cells.
To further understand the functional mechanisms behind Nr-CWS induced NK cell-mediated anti-tumor responses, the functional markers of NK cell playing essential roles in immuno-regulation, antiviral activity, and immune-surveillance were analyzed. As well known, activated NK cells express certain markers like CD69 [26] and functional molecules such as TRAIL and FasL, via which NK cells kill targets through apoptotic pathways [27, 28]. Our data indicates that Nr-CWS increased the pro-apoptotic molecules TRAIL and FasL expression of NK cells in vivo (Figure 2A, Supplementary Figure S2A).
To further explore related mechanisms, the purified or sorted NK cells treated with Nr-CWS were assessed by determining the expression of phenotype markers and cytotoxic molecules, production of intracellular and secreted cytokines, and cytotoxicity against tumor cells. The data revealed that the CD69, TRAIL, and FasL expression were strikingly up-regulated after 24 h treatment with Nr-CWS (Figures 2C,D, Supplementary Figures S2B,C), which is consistent with in vivo results. In addition to TRAIL and FasL, NK cells may exert their cytotoxic function directly through GrzB and Prf, which can perforate cell membranes and directly kill target cells [29]. Consistently, the suitable amount of Nr-CWS can significantly increase the production of GrzB and Prf of NK cells after 24 and 48 h cell culture (Figures 3A,B, Supplementary Figures S3A,B). NK cells were defined as CD27lowCD11blow, CD27highCD11blow, CD27highCD11bhigh, and CD27lowCD11bhigh four subgroups by CD27 and CD11b expression. The CD27high NK cell subgroups display the highest secretion of cytokines, while the CD27lowCD11bhigh subgroup is the terminally differentiated cells that have the strongest cytotoxic capacity [30–32]. Our data indicated that Nr-CWS decreased CD27 expression after treatment for 24 or 48 h (Figures 3C,D, Supplementary Figures S3C,D), suggesting that Nr-CWS could enhance NK cell function through promoting NK cell into CD27lowCD11bhigh terminally differentiated subgroup.
Besides their direct effect on target, NK cells, particularly the terminally differentiated ones, can regulate anti-tumor or anti-pathogen responses through secretion of different cytokines, chemokines, and growth factors [14, 33, 34], which have been related to NK cell terminal differentiation [35]. In order to get deeper insight into Nr-CWS induced NK cells maturation and cytokine production, we measured the levels of cytokines in the serum and supernatants. In both serum and the supernatants, the levels of IFN-γ and TNF-α were strikingly up-regulated after Nr-CWS treatment (Figure 4, Supplementary Figure S4), suggesting that Nr-CWS could enhance anti-tumor activities of NK cells through regulating cytokines production.
Finally, we also tested the cytotoxicity in vitro using sorted NK cells. The result indicated that Nr-CWS could augment cytotoxic activity of NK cells (Figure 5). In light of the cytokine analysis data (Figures 4D,E), it is reasonable to believe that Nr-CWS may enhance the anti-tumor activity and augmentation of NK cell functions. Taken together, our study revealed a new profile of NK cells activation by Nr-CWS and provided a new mode of action of Nr-CWS as a new immune modulator in cancer therapy. Although we understand that Nr-CWS promotes NK cell function and maturation, it still needs to study its underlying mechanisms. In future experiments, in order to elucidate its underlying mechanism of action, it may be necessary to identify Nr-CWS-binding receptors on NK cells, as well as intracellular signaling pathways.
Conclusion
In conclusion, our study demonstrated that Nr-CWS could suppress the lung metastasis of B16F10 melanoma model, which may be through its effect on NK cells. The possible mechanisms include: up-regulation of the expression of FasL and TRAIL, promotion of the production of cytotoxic cytokines (IFN-γ, TNF-α) and granules (Prf, GrzB), promotion of NK cell differentiation into the terminally matured CD27low CD11bhigh subgroup and enhancement of the cytotoxic activity of NK cell.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Ethics Statement
The animal study was reviewed and approved by the ethics committee of Jinzhou Medical University.
Author Contributions
JW, BH, XZ, and GW were involved in the design of the study. JW, BH, MM, XH, HeD, and YL performed the experiments and analyzed the data. JW, HoD, XZ, and GW contributed to the writing and editing of the manuscript.
Funding
The research was supported by the Scientific Research Fund of Liaoning Provincial Education Department under Grant JYTJCZR2020053.
Conflict of Interest
Authors HeD and YL are employed by Greatest Biopharma Limited Company.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.por-journal.com/articles/10.3389/pore.2022.1610555/full#supplementary-material
References
1. Kuyukina, MS, Ivshina, IB, Baeva, TA, Kochina, OA, Gein, SV, and Chereshnev, VA. Trehalolipid Biosurfactants from Nonpathogenic Rhodococcus Actinobacteria with Diverse Immunomodulatory Activities. N Biotechnol (2015) 32:559–68. doi:10.1016/j.nbt.2015.03.006
2. Azuma, I, Taniyama, T, Yamawaki, M, Sugimura, K, and Yamamura, Y. Adjuvant and Antitumor Activities of Nocardia Cell-wall Skeletons. Gan (1976) 67:733–6.
3. Meng, Y, Sun, J, Wang, X, Ma, Y, Kong, C, Zhang, G, et al. The Biological Macromolecule Nocardia Rubra Cell-wall Skeleton as an Avenue for Cell-Based Immunotherapy. J Cel Physiol (2019) 2019. doi:10.1002/jcp.28182
4. Wang, G, Wu, J, Miao, M, Dou, H, Nan, N, Shi, M, et al. Nocardia Rubra Cell-wall Skeleton Promotes CD4(+) T Cell Activation and Drives Th1 Immune Response. Int J Biol Macromol (2017) 101:398–407. doi:10.1016/j.ijbiomac.2017.03.060
5. Tao, Y, Wang, G, Zhai, J, and Zhang, N. Functional Modulation of CD8+ T Cell by Approved Novel Immune Enhancer: Nocardia Rubra Cell-Wall Skeletons (Nr-CWS). Int Immunopharmacol (2020) 78:106023. doi:10.1016/j.intimp.2019.106023
6. Chen, W, Zhang, Y, Zhao, C, Shao, S, Zhang, Y, Li, X, et al. Nocardia Rubra Cell Wall Skeleton Up-Regulates T Cell Subsets and Inhibits PD-1/pd-L1 Pathway to Promote Local Immune Status of Patients with High-Risk Human Papillomavirus Infection and Cervical Intraepithelial Neoplasia. Front Immunol (2020) 11:612547. doi:10.3389/fimmu.2020.612547
7. Wang, Y, Hu, Y, Ma, B, Wang, F, Liu, S, Xu, J, et al. Nocardia Rubra Cell wall Skeleton Accelerates Cutaneous Wound Healing by Enhancing Macrophage Activation and Angiogenesis. J Int Med Res (2018) 46:2398–409. doi:10.1177/0300060518764210
8. Rohde, D, Gastl, G, Biesterfeld, S, Plante, M, and Jakse, G. Local Expression of Cytokines in Rat Bladder Carcinoma Tissue after Intravesical Treatment with Nocardia Rubra Cell wall Skeleton and Bacille-Calmette-Guerin. Urol Res (1997) 25:19–24. doi:10.1007/BF00941901
9. Huang, C, Tang, X, Li, S, Wang, Q, Xie, B, Xu, J, et al. Immunopotentiator Aikejia Improves the Therapeutic Efficacy of PD-1/pd-L1 Immunosuppressive Pathway in CT26.WT Cancer Cell. J Cancer (2019) 10:3472–80. doi:10.7150/jca.29672
10. Dell'Oste, V, Biolatti, M, Galitska, G, Griffante, G, Gugliesi, F, Pasquero, S, et al. Tuning the Orchestra: HCMV vs. Innate Immunity. Front Microbiol (2020) 11:661. doi:10.3389/fmicb.2020.00661
11. Liu, S, Galat, V, Galat, Y, Lee, YKA, Wainwright, D, and Wu, J. NK Cell-Based Cancer Immunotherapy: from Basic Biology to Clinical Development. J Hematol Oncol (2021) 14:7. doi:10.1186/s13045-020-01014-w
12. Zhang, X, and Wei, H. Role of Decidual Natural Killer Cells in Human Pregnancy and Related Pregnancy Complications. Front Immunol (2021) 12:728291. doi:10.3389/fimmu.2021.728291
13. Yu, G, Xu, X, Vu, MD, Kilpatrick, ED, and Li, XC. NK Cells Promote Transplant Tolerance by Killing Donor Antigen-Presenting Cells. J Exp Med (2006) 203:1851–8. doi:10.1084/jem.20060603
14. Morvan, MG, and Lanier, LL. NK Cells and Cancer: You Can Teach Innate Cells New Tricks. Nat Rev Cancer (2016) 16:7–19. doi:10.1038/nrc.2015.5
15. Grudzien, M, and Rapak, A. Effect of Natural Compounds on NK Cell Activation. J Immunol Res (2018) 2018:4868417. doi:10.1155/2018/4868417
16. Bi, J, and Wang, X. Molecular Regulation of NK Cell Maturation. Front Immunol (2020) 11:1945. doi:10.3389/fimmu.2020.01945
17. Debska-Zielkowska, J, Moszkowska, G, Zielinski, M, Zielinska, H, Dukat-Mazurek, A, Trzonkowski, P, et al. KIR Receptors as Key Regulators of NK Cells Activity in Health and Disease. Cells (2021) 10:1777. doi:10.3390/cells10071777
18. Wu, F, Ding, XY, Li, XH, Gong, MJ, An, JQ, and Huang, SL. Correlation between Elevated Inflammatory Cytokines of Spleen and Spleen index in Acute Spinal Cord Injury. J Neuroimmunol (2020) 344:577264. doi:10.1016/j.jneuroim.2020.577264
19. Chen, W, Liu, J, Meng, J, Lu, C, Li, X, Wang, E, et al. Macrophage Polarization Induced by Neuropeptide Methionine Enkephalin (MENK) Promotes Tumoricidal Responses. Cancer Immunol Immunother (2012) 61:1755–68. doi:10.1007/s00262-012-1240-6
20. Wu, X, Cheng, B, Cai, ZD, and Lou, LM. Determination of the Apoptotic index in Osteosarcoma Tissue and its Relationship with Patients Prognosis. Cancer Cel Int (2013) 13:56. doi:10.1186/1475-2867-13-56
21. Sivori, S, Carlomagno, S, Pesce, S, Moretta, A, Vitale, M, and Marcenaro, E. TLR/NCR/KIR: Which One to Use and when? Front Immunol (2014) 5:105. doi:10.3389/fimmu.2014.00105
22. Kim, S, Iizuka, K, Aguila, HL, Weissman, IL, and Yokoyama, WM. In Vivo natural Killer Cell Activities Revealed by Natural Killer Cell-Deficient Mice. Proc Natl Acad Sci U S A (2000) 97:2731–6. doi:10.1073/pnas.050588297
23. Street, SE, Zerafa, N, Iezzi, M, Westwood, JA, Stagg, J, Musiani, P, et al. Host Perforin Reduces Tumor Number but Does Not Increase Survival in Oncogene-Driven Mammary Adenocarcinoma. Cancer Res (2007) 67:5454–60. doi:10.1158/0008-5472.CAN-06-4084
24. Krneta, T, Gillgrass, A, Chew, M, and Ashkar, AA. The Breast Tumor Microenvironment Alters the Phenotype and Function of Natural Killer Cells. Cell Mol Immunol (2016) 13:628–39. doi:10.1038/cmi.2015.42
25. Eckelhart, E, Warsch, W, Zebedin, E, Simma, O, Stoiber, D, Kolbe, T, et al. A Novel Ncr1-Cre Mouse Reveals the Essential Role of STAT5 for NK-Cell Survival and Development. Blood (2011) 117:1565–73. doi:10.1182/blood-2010-06-291633
26. Bjorkstrom, NK, Strunz, B, and Ljunggren, HG. Natural Killer Cells in Antiviral Immunity. Nat Rev Immunol (2021) 22:112–23. doi:10.1038/s41577-021-00558-3
27. Sag, D, Ayyildiz, ZO, Gunalp, S, and Wingender, G. The Role of TRAIL/DRs in the Modulation of Immune Cells and Responses. Cancers (Basel) (2019) 11:1469. doi:10.3390/cancers11101469
28. Rossin, A, Miloro, G, and Hueber, AO. TRAIL and FasL Functions in Cancer and Autoimmune Diseases: Towards an Increasing Complexity. Cancers (Basel) (2019) 11:639. doi:10.3390/cancers11050639
29. Fehniger, TA, Cai, SF, Cao, X, Bredemeyer, AJ, Presti, RM, French, AR, et al. Acquisition of Murine NK Cell Cytotoxicity Requires the Translation of a Pre-existing Pool of Granzyme B and Perforin mRNAs. Immunity (2007) 26:798–811. doi:10.1016/j.immuni.2007.04.010
30. Abel, AM, Yang, C, Thakar, MS, and Malarkannan, S. Natural Killer Cells: Development, Maturation, and Clinical Utilization. Front Immunol (2018) 9:1869. doi:10.3389/fimmu.2018.01869
31. Fu, B, Wang, F, Sun, R, Ling, B, Tian, Z, and Wei, H. CD11b and CD27 Reflect Distinct Population and Functional Specialization in Human Natural Killer Cells. Immunology (2011) 133:350–9. doi:10.1111/j.1365-2567.2011.03446.x
32. Chiossone, L, Chaix, J, Fuseri, N, Roth, C, Vivier, E, and Walzer, T. Maturation of Mouse NK Cells Is a 4-stage Developmental Program. Blood (2009) 113:5488–96. doi:10.1182/blood-2008-10-187179
33. Bald, T, Krummel, MF, Smyth, MJ, and Barry, KC. The NK Cell-Cancer Cycle: Advances and New Challenges in NK Cell-Based Immunotherapies. Nat Immunol (2020) 21:835–47. doi:10.1038/s41590-020-0728-z
34. Vivier, E, Raulet, DH, Moretta, A, Caligiuri, MA, Zitvogel, L, Lanier, LL, et al. Innate or Adaptive Immunity? the Example of Natural Killer Cells. Science (2011) 331:44–9. doi:10.1126/science.1198687
Keywords: cancer, metastasis, natural killer cell, Nocardia rubra cell-wall skeleton, cytotoxicity
Citation: Wu J, He B, Miao M, Han X, Dai H, Dou H, Li Y, Zhang X and Wang G (2022) Enhancing Natural Killer Cell-Mediated Cancer Immunotherapy by the Biological Macromolecule Nocardia rubra Cell-Wall Skeleton. Pathol. Oncol. Res. 28:1610555. doi: 10.3389/pore.2022.1610555
Received: 26 April 2022; Accepted: 17 August 2022;
Published: 30 August 2022.
Edited by:
Andrea Ladányi, National Institute of Oncology (NIO), HungaryCopyright © 2022 Wu, He, Miao, Han, Dai, Dou, Li, Zhang and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaoqing Zhang, emhhbmd4aWFvcWluZzIwMTRAMTI2LmNvbQ==; Guangchuan Wang, Y2h1YW41NjBAMTYzLmNvbQ==